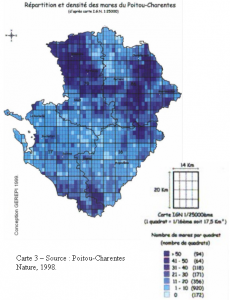
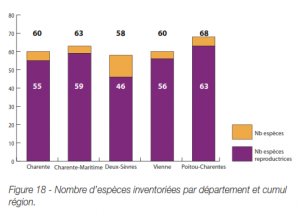
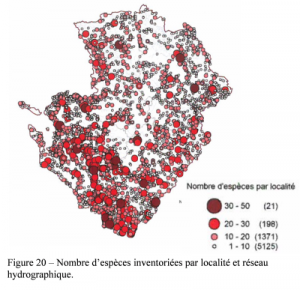
La durée de l’incubation des œufs est assez variable en fonction des conditions environnementales, que ce soit d’une espèce à l’autre et au sein d’une même espèce. Certaines libellules, adaptées aux conditions temporaires, peuvent éclore quelques jours seulement après la ponte. C’est notamment le cas du Sympétrum strié Sympetrum striolatum chez qui les premières éclosions ont été observées 14 jours seulement après la ponte (Jourde, inédit).
Un environnement défavorable peut entraîner l’arrêt du développement de l’œuf, qui entre en diapause. Cette stratégie adaptative permet aux œufs de passer l’hiver dans un état de dormance. L’éclosion retardée permet aux fragiles larves de commencer leur croissance alors que l’eau se réchauffe et que les proies se font plus abondantes. La période d’incubation peut dès lors se compter en mois.
La proportion d’œufs entrant en diapause durant l’hiver augmente à mesure que la ponte est tardive. Chez certaines espèces, au sein d’une même génération, certains œufs entrent en diapause, d’autres pas ; une façon de ne pas mettre tous ses œufs dans le même panier ?
L’apparence des œufs se modifie sensiblement au fil du développement embryonnaire. Blancs à jaune pâle après la ponte, ils s’assombrissent rapidement. Après quelques jours, il est possible d’observer par transparence le développement de l’embryon.
Éclosion et développement larvaire
Chez les espèces à diapause hivernale, l’éclosion se fait de façon relativement synchrone à la fin de l’hiver. Les espèces qui ne développent pas de diapause, ou chez qui une partie seulement de la population entre en dormance, ont une période d’éclosion beaucoup plus étalée.
Les larves de quelques espèces tropicales peuvent se développer dans l’humidité de la litière du sol mais en Europe, toutes vivent dans l’eau. La période de développement larvaire est très variable d’une espèce à l’autre. Elle est essentiellement conditionnée par la température de l’eau et la disponibilité en proies. Pour faire simple, plus l’eau est chaude et riche en nutriments, plus les larves se développent rapidement. Plus elle est pauvre et froide, moins vite les larves atteindront leur dernier stade de croissance.
Dans la nature, on observe donc de grandes différences entre espèces mais aussi au sein des espèces selon les endroits où elles vivent. La larve de l’Aeschne affine Aeshna affinis peut se développer en 4 mois dans les milieux temporaires saumâtres de Charente-Maritime. Dans les sources froides, celle du Cordulégastre annelé Cordulegaster boltonii met habituellement 3 à 4 ans pour devenir adulte. Cette faculté d’adaptation permet aux libellules de coloniser la quasi-totalité des milieux aquatiques.

La croissance des larves se fait par une succession de 8 à 18 mues, généralement de 11 à 13. Le premier stade est bref. Une prolarve vermiforme émerge de l’œuf et se transforme rapidement, parfois dès l’éclosion, en une larve pourvue de pattes, d’antennes et d’un masque mentonnier. A partir de là, les mues s’enchaînent. Au fil de son développement, la larve gagne en taille et en complexité pour finalement aboutir à un insecte prêt à quitter le milieu aquatique et à conquérir les airs.
Chez certaines espèces comme le Sympétrum méridional Sympetrum méridionale, les derniers jours de développement larvaire peuvent s’opérer dans des points d’eau asséchés. Les larves se maintiennent à l’abri sous des algues humides ou dans les touffes de végétaux.


Mode de déplacement
Si la prolarve n’est capable de se mouvoir qu’en se tortillant à la façon d’un asticot, les larves sont pourvues de pattes fonctionnelles, qui leur permettent de se déplacer dans la végétation aquatique, les enchevêtrements de racines, les amas de végétaux ou les sédiments du fond.
Les zygoptères peuvent nager en agitant leur abdomen. Les lamelles caudales servent alors de godille. Pour fuir, mais aussi parfois pour attaquer, les larves d’anisoptères chassent violemment et de façon répétée l’eau contenue dans leur ampoule rectale. Elles se propulsent par réaction.
Nourriture et chasse
Les larves sont carnassières et se nourrissent d’animaux vivants qu’elles repèrent à vue mais aussi vraisemblablement par des poils sensibles aux vibrations engendrées par le déplacement des proies. Selon leur stade de développement, elles peuvent capturer des proies de taille très variable. Il s’agit généralement d’animalcules durant les premiers stades de croissance mais les grandes espèces peuvent s’en prendre exceptionnellement à des tritons ou des alevins à la fin de leur vie larvaire. L’essentiel du régime est composé de petits crustacés (cladocères, gammares) et de larves d’insectes, dont les chironomes et les moustiques constituent une part importante. Elles consomment aussi les larves d’autres espèces de libellules, voire les premiers stades de leur propre espèce.
Les proies sont chassées à l’approche ou à l’affût. Dans le premier cas, les larves arpentent doucement la végétation immergée ou accumulée sur le fond et débusquent leur proie au détour d’une feuille ou d’une brindille. Dans le second cas, la larve immobile et généralement parfaitement camouflée dans son environnement, souvent même partiellement enfouie dans les sédiments, attend le passage d’une proie. Quand cette dernière est suffisamment proche, la larve projette une sorte de bras articulé situé sous la tête et muni à son extrémité de deux crochets préhensiles : le masque mentonnier ou mentum. C’est la soudaine compression du corps de la larve et la mobilité de ses fluides corporels qui permettent l’extension du masque à la façon d’une langue de belle-mère.
Philippe Jourde
Bibliographie
Jourde P., 2000 – Nouvelles données de captures d’odonates par un végétal non carnivore. Martinia, 16 (1) 3-7.