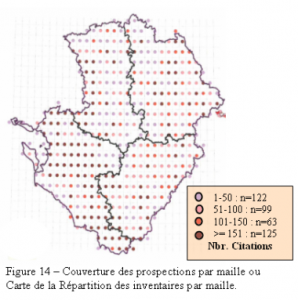
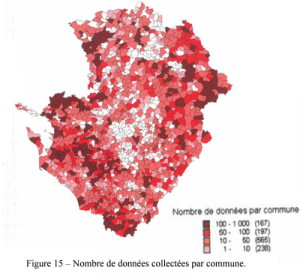
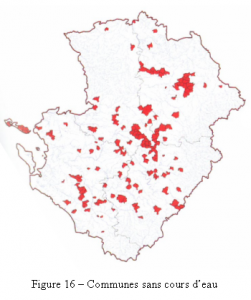
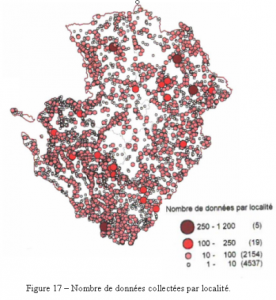
Les sources
 Localisées en tête de bassin, les sources forment un habitat aquatique spécifique qui correspond à la réunion du sous-sol, du milieu aquatique et du milieu aérien. La biodiversité y est le plus souvent pauvre, mais composée d’espèces spécifiques (mollusques, crustacés et autres invertébrés). Les sources sont généralement des milieux stables sur le plan physico-chimique notamment si l’eau provient de nappes phréatiques profondes, même si les pollutions diffuses sont de plus en plus importantes. La température de l’eau est fraîche et stable. En Poitou-Charentes, ces sources sont encore nombreuses et se déversent souvent dans des bassins, qui débordent ensuite pour former des ruisselets. Elles sont les milieux de prédilection pour la reproduction du Cordulégastre annelé Cordulegaster boltonii dont les larves supportent l’absence de végétation immergée et la fraicheur de l’eau. Si la végétation aquatique (hélophytes et hydrophytes) s’y développe, d’autres espèces comme le Calopteryx vierge Calopteryx virgo ou l’Agrion de Mercure Coenagrion mercuriale peuvent aussi occuper ces milieux.
Localisées en tête de bassin, les sources forment un habitat aquatique spécifique qui correspond à la réunion du sous-sol, du milieu aquatique et du milieu aérien. La biodiversité y est le plus souvent pauvre, mais composée d’espèces spécifiques (mollusques, crustacés et autres invertébrés). Les sources sont généralement des milieux stables sur le plan physico-chimique notamment si l’eau provient de nappes phréatiques profondes, même si les pollutions diffuses sont de plus en plus importantes. La température de l’eau est fraîche et stable. En Poitou-Charentes, ces sources sont encore nombreuses et se déversent souvent dans des bassins, qui débordent ensuite pour former des ruisselets. Elles sont les milieux de prédilection pour la reproduction du Cordulégastre annelé Cordulegaster boltonii dont les larves supportent l’absence de végétation immergée et la fraicheur de l’eau. Si la végétation aquatique (hélophytes et hydrophytes) s’y développe, d’autres espèces comme le Calopteryx vierge Calopteryx virgo ou l’Agrion de Mercure Coenagrion mercuriale peuvent aussi occuper ces milieux.
Dans la région, ces milieux sont le plus souvent de faible superficie. Quelques sites caractéristiques d’importance peuvent être cités : les sources du Thouet (79), de la Lèche (16), la fontaine romaine de Gros-Roc (Le Douhet,17) ou encore les sources de la Luire et du Glandon (86).
Les suintements
 Les suintements se caractérisent par un écoulement d’eau très faible sur des substrats vaseux ou boueux. Ces biotopes naturels de petite taille sont généralement localisés dans des prairies, mais on peut les rencontrer dans bien d’autres milieux comme les carrières ou autour d’étangs (fuite située en aval d’ouvrage par exemple) et ils peuvent donc être à cette occasion d’origine anthropique. Sur ces milieux originaux, seules les espèces dont les larves survivent dans très peu d’eau, parfois même dans un sol juste maintenu humide, se développent. Les Orthétrums brun Orthetrum brunneum et bleuissant O. coerulescens affectionnent ces habitats lorsqu’ils sont bien ensoleillés, le premier préférant les filets d’eau suintants en milieu dénudé alors que la présence du second nécessite plus de végétation. C. boltonii peut compléter ce mince cortège, car sa biologie, notamment la capacité à pondre directement dans les sédiments, est très adaptée aux suintements. De même l’Agrion nain Ischnura pumilio, espèce particulièrement pionnière, pourra apparaître et trouver dans les suintements les conditions favorables à son développement, notamment si la profondeur de la nappe d’eau augmente légèrement de place en place. Si ces suintements sont très ombragés, on ne retrouvera plus que le cordulégastre, mais lorsqu’ils sont ensoleillés et permettent le développement de la végétation aquatique, il arrive d’y rencontrer C. mercuriale, C. Tenelenum et P. nymphula.
Les suintements se caractérisent par un écoulement d’eau très faible sur des substrats vaseux ou boueux. Ces biotopes naturels de petite taille sont généralement localisés dans des prairies, mais on peut les rencontrer dans bien d’autres milieux comme les carrières ou autour d’étangs (fuite située en aval d’ouvrage par exemple) et ils peuvent donc être à cette occasion d’origine anthropique. Sur ces milieux originaux, seules les espèces dont les larves survivent dans très peu d’eau, parfois même dans un sol juste maintenu humide, se développent. Les Orthétrums brun Orthetrum brunneum et bleuissant O. coerulescens affectionnent ces habitats lorsqu’ils sont bien ensoleillés, le premier préférant les filets d’eau suintants en milieu dénudé alors que la présence du second nécessite plus de végétation. C. boltonii peut compléter ce mince cortège, car sa biologie, notamment la capacité à pondre directement dans les sédiments, est très adaptée aux suintements. De même l’Agrion nain Ischnura pumilio, espèce particulièrement pionnière, pourra apparaître et trouver dans les suintements les conditions favorables à son développement, notamment si la profondeur de la nappe d’eau augmente légèrement de place en place. Si ces suintements sont très ombragés, on ne retrouvera plus que le cordulégastre, mais lorsqu’ils sont ensoleillés et permettent le développement de la végétation aquatique, il arrive d’y rencontrer C. mercuriale, C. Tenelenum et P. nymphula.
Les ruisselets et ruisseaux en milieu boisé
Il s’agit de petits cours d’eau de tête de bassins alimentés par les sources et ayant une faible largeur. Le courant est le plus souvent vif générant une eau oxygénée, même si parfois les débits ne sont pas très importants (quelques litres par seconde).
 Les ruisseaux traversant des zones boisées sont pauvres en hydrophytes et hélophytes. Le cortège odonatologique est alors limité. On trouve souvent C. boltoni sur l’amont. Sur les parties aval et un peu plus larges, d’autres anisoptères font leur apparition : l’Aeschne paisible Boyera irene, le Gomphe à crochets Onychogomphus uncatus et le Gomphe vulgaire Gomphus vulgatissimus, profitent des eaux rapides et bien oxygénées dans lesquelles leurs larves trouvent des conditions optimales de développement ; si le courant est plus lent, on peut parfois rencontrer la Cordulie métallique Somatochlora metallica dont les femelles cherchent souvent la protection de la végétation rivulaire et l’ombre pour pondre. Quant aux zygoptères, C. virgo et quelques espèces ubiquistes (l’Agrion à large pattes Platycnemis pennipes par exemple) peuvent se cantonner sur les rares portions où la strate arborée est traversée par le soleil.
Les ruisseaux traversant des zones boisées sont pauvres en hydrophytes et hélophytes. Le cortège odonatologique est alors limité. On trouve souvent C. boltoni sur l’amont. Sur les parties aval et un peu plus larges, d’autres anisoptères font leur apparition : l’Aeschne paisible Boyera irene, le Gomphe à crochets Onychogomphus uncatus et le Gomphe vulgaire Gomphus vulgatissimus, profitent des eaux rapides et bien oxygénées dans lesquelles leurs larves trouvent des conditions optimales de développement ; si le courant est plus lent, on peut parfois rencontrer la Cordulie métallique Somatochlora metallica dont les femelles cherchent souvent la protection de la végétation rivulaire et l’ombre pour pondre. Quant aux zygoptères, C. virgo et quelques espèces ubiquistes (l’Agrion à large pattes Platycnemis pennipes par exemple) peuvent se cantonner sur les rares portions où la strate arborée est traversée par le soleil.
Parmi les ruisseaux picto-charentais abritant ce cortège on peut citer : le ruisseau de la Feuillante (Ligugé, 86), le ruisseau de la Lizonne (Bioussac, 16), le ruisseau du Pressoir (79), le Bruant à Saint-Porchaire (17).
Ruisseaux et rivières peu profondes en milieu ouvert
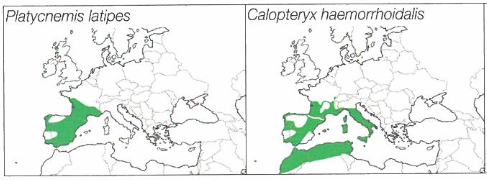
 Sur les petits cours d’eau coulant en milieu ouvert (prairies), la végétation aquatique (hydrophytes) et celle des berges (hélophytes) sont généralement riches, ce qui est favorable à l’installation de cortèges odonatologiques beaucoup plus diversifiés. La nature du fond (sableux, graveleux, vaseux…), la densité de la ripisylve et surtout la vitesse du courant, conditionnent la présence des différentes espèces en fonction des exigences de leurs larves. En ce qui concerne les anisoptères, parmi les espèces principales, le Gomphe vulgaire Gomphus vulgatissimus est le premier à apparaître en saison. Il est remplacé ensuite par O. uncatus dans les parties vives et le Gomphe à pinces Onychogomphus forcipatus sur les portions plus calmes. Dans les secteurs très ensoleillés, la Libellule fauve Libellula fulva est aussi caractéristique de ces cours d’eau. Plus tard en saison, on peut rencontrer sur les mêmes stations O. brunneum et O. coerulescens. Si les berges sont plantées d’arbres offrant des systèmes racinaires bien fournis, les populations de B. irene à la recherche de recoins sombres pour la ponte et la vie larvaire, peuvent prospérer. Comme zygoptères, C. mercuriale et C. virgo, qui apprécient les eaux bien oxygénées et plutôt rapides, sont presque omniprésents dans ces habitats, tant qu’ils peuvent bénéficier d’herbiers en abondance (Callitriche spp, Apium nodiflorum, …) pour servir de supports de ponte puis d’abri aux larves. Au sud de la région, ils sont rejoints par le Caloptéryx hémorrhoïdal Calopteryx haemorrhoidalis. Dans les zones à courant plus modéré de ces ruisseaux et petites rivières, on peut parfois observer des espèces davantage inféodées aux milieux stagnants ou aux parties calmes des milieux courants comme la Cordulie au corps fin Oxygastra curtisii, l’Agrion jouvencelle Coenagrion puella ou l’Agrion à larges pattes Platycnemis pennipes et le Caloptéryx éclatant Calopteryx spendens. Les larves de ces espèces s’accommodent voire préfèrent des fonds vaseux aux substrats uniquement graveleux.
Sur les petits cours d’eau coulant en milieu ouvert (prairies), la végétation aquatique (hydrophytes) et celle des berges (hélophytes) sont généralement riches, ce qui est favorable à l’installation de cortèges odonatologiques beaucoup plus diversifiés. La nature du fond (sableux, graveleux, vaseux…), la densité de la ripisylve et surtout la vitesse du courant, conditionnent la présence des différentes espèces en fonction des exigences de leurs larves. En ce qui concerne les anisoptères, parmi les espèces principales, le Gomphe vulgaire Gomphus vulgatissimus est le premier à apparaître en saison. Il est remplacé ensuite par O. uncatus dans les parties vives et le Gomphe à pinces Onychogomphus forcipatus sur les portions plus calmes. Dans les secteurs très ensoleillés, la Libellule fauve Libellula fulva est aussi caractéristique de ces cours d’eau. Plus tard en saison, on peut rencontrer sur les mêmes stations O. brunneum et O. coerulescens. Si les berges sont plantées d’arbres offrant des systèmes racinaires bien fournis, les populations de B. irene à la recherche de recoins sombres pour la ponte et la vie larvaire, peuvent prospérer. Comme zygoptères, C. mercuriale et C. virgo, qui apprécient les eaux bien oxygénées et plutôt rapides, sont presque omniprésents dans ces habitats, tant qu’ils peuvent bénéficier d’herbiers en abondance (Callitriche spp, Apium nodiflorum, …) pour servir de supports de ponte puis d’abri aux larves. Au sud de la région, ils sont rejoints par le Caloptéryx hémorrhoïdal Calopteryx haemorrhoidalis. Dans les zones à courant plus modéré de ces ruisseaux et petites rivières, on peut parfois observer des espèces davantage inféodées aux milieux stagnants ou aux parties calmes des milieux courants comme la Cordulie au corps fin Oxygastra curtisii, l’Agrion jouvencelle Coenagrion puella ou l’Agrion à larges pattes Platycnemis pennipes et le Caloptéryx éclatant Calopteryx spendens. Les larves de ces espèces s’accommodent voire préfèrent des fonds vaseux aux substrats uniquement graveleux.
Le ruisseau du Miosson (Nieuil-l’Espoir, 86), le ruisseau de la Luire (Lésigny, 86), le ruisseau de l’Or (Champagne-Mouton, 16) sont des exemples de ruisseaux ouverts dans la région. La Boivre (Biard, 86), la Boëme (Mouthiers-sur-Boëme, 16) sont, quant à elles, des exemples typiques de petites rivières. La Seugne (de Jonzac à Les Gonds, 17) présente un cortège odonatologique à la fois caractéristique et abondant où alternent espaces ouverts et fermés.
Les grandes rivières rapides et les secteurs d’accélération des grands cours d’eau
Il s’agit de cours d’eau ou de portions de cours d’eau (zones ponctuelles d’accélération du courant) d’une largeur relativement grande correspondant souvent à la « zone à barbeaux ». Leur profondeur est très variable et leur morphologie fait alterner zones profondes où l’eau peut parfois circuler entre de gros blocs de pierre, plats courants et radiers peu profonds où l’eau court généralement sur un lit de graviers. Sur ces rivières, on trouve fréquemment de grands herbiers de renoncules aquatiques. En Poitou-Charentes, c’est dans ces milieux lotiques que l’on trouve les cortèges dominés par les gomphidés qui occupent, successivement dans la saison, les mêmes niches écologiques : G. vulgatissimus puis le Gomphe semblable Gomphus simillimus, O. uncatus et O. forcipatus. Tous ces gomphes ont également une phénologie bien distincte et les pics d’émergence puis d’activité se succèdent dans la saison sur un même cours d’eau (figure 1). La qualité du substrat est un facteur limitant important en ce qui concerne le développement des larves de ces espèces. O. uncatus et G. simillimus ont besoin de sable, de graviers, voire de gros cailloux, les portions de rivières sur lesquelles le limon est très important et où la vase domine sont en revanche désertés. Au contraire, les larves de G. vulgatissimus et d’O. forcipatus acceptent fort bien les sols vaseux dans lesquels elles s’enfouissent pour chasser. Le Gomphe à pinces, de ce fait, sera surtout présent au niveau des retours de courant, des remous, à l’intérieur des méandres.
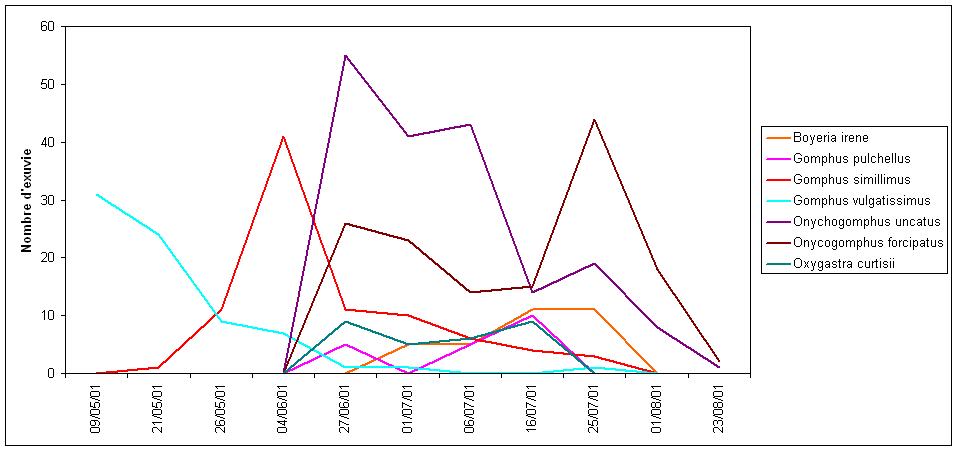
Dans la Vienne uniquement, et plus rarement, on peut aussi trouver le rare Gomphe à pattes jaunes Gomphus flavipes. D’autres espèces d’anisoptères accompagnent régulièrement les gomphidés comme B. irene, O. curtisii. Quant aux zygoptères, E. lindenii et le Caloptéryx éclatant C. splendens sont les plus caractéristiques mais, ponctuellement, de nombreuses autres espèces peuvent être observées au gré de la présence de micro-habitats plus ou moins lentiques : C. puella, L. fulva, P. pennipes …
 Dans le Poitou-Charentes, la Gartempe (86), la Creuse (86), la Dronne (16, 17), le cours moyen de la Charente (16, 86) offrent plus qu’ailleurs ce type d’habitat. Toutefois, toutes les rivières moyennes et grandes de la région peuvent posséder des secteurs où le courant s’accélère : radiers, gués, aval immédiat de barrages ou de digues.
Dans le Poitou-Charentes, la Gartempe (86), la Creuse (86), la Dronne (16, 17), le cours moyen de la Charente (16, 86) offrent plus qu’ailleurs ce type d’habitat. Toutefois, toutes les rivières moyennes et grandes de la région peuvent posséder des secteurs où le courant s’accélère : radiers, gués, aval immédiat de barrages ou de digues.
Les zones calmes des rivières et grands cours d’eau
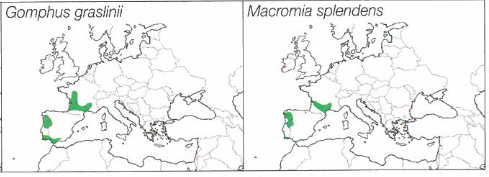
 Ces secteurs où le courant est très ralenti peuvent se rencontrer sur des cours d’eau de toutes tailles. Sur ceux d’une largeur moyenne à grande, il s’agit de la « zone à brème » localisée sur la partie aval du cours. Les cortèges odonatologiques sont riches et beaucoup d’espèces profitent des eaux réchauffées l’été, des fonds riches en sédiments et en débris végétaux ainsi que de la forte végétalisation du cours d’eau et de ses rives. Ailleurs, sur les rivières plus rapides et souvent de dimensions plus modestes, de nombreux ouvrages hydrauliques (barrages, seuils) ont souvent modifié de manière importante le régime naturel du cours d’eau, et des zones calmes, relativement profondes avec des fonds vaseux plantés de nombreux hydrophytes, ont permis aux mêmes cortèges de s’installer. L’espèce caractéristique qui domine ici et qui survole inlassablement ces eaux calmes est l’Anax empereur Anax imperator parfois remplacé par l’Anax napolitain Anax parthenope, qui moins agressif, supporte souvent mal la concurrence avec le premier. Tandis que L. fulva puis O. curtisii, plus tard en saison, patrouillent le long des berges, le Gomphe de Graslin Gomphus graslinii demeure au-dessus des herbiers flottants. B. irene peut profiter des berges les plus boisées. Sur certains secteurs de la Charente et de la Dronne, on peut même rencontrer la très rare Cordulie splendide Macromia splendens. Le cortège caractéristique est aussi composé de nombreuses espèces de zygoptères. Des populations nombreuses de C. splendens sont généralement présentes. Au sud de la région, elles côtoient et parfois se mélangent avec les peuplements du Caloptéryx occitan Calopteryx xanthostoma. Les zones enherbées des rives servent de terrain de chasse aux Platycnémididés, surtout l’Agrion blanchâtre Platycnemis latipes et P. pennipes alors que les nombreux herbiers d’hydrophytes affleurant (Myriophyllum spp. et Ceratophyllum spp. surtout) leur permettent de déposer leur ponte puis d’abriter les larves. Sur l’eau, au-dessus de ces mêmes herbiers, E. lindenii est omniprésent alors que les deux autres espèces d’Erythromma, la Naïade au corps vert E. viridulum et plus rarement la Naïade aux yeux rouges E. najas, marquent une préférence pour les hydrophytes flottants (Nympha lutea ou Lemna spp. notamment). Bien sûr, plusieurs espèces plus ubiquistes ou ayant une écologie plus plastique enrichissent souvent ces cortèges, en particulier C. puella, I. elegans, l’Agrion orangé Platycnemis acutipennis, le Leste vert Chalcolestes viridis, le Gomphe joli Gomphus pulchellus, ou encore O. forcipatus.
Ces secteurs où le courant est très ralenti peuvent se rencontrer sur des cours d’eau de toutes tailles. Sur ceux d’une largeur moyenne à grande, il s’agit de la « zone à brème » localisée sur la partie aval du cours. Les cortèges odonatologiques sont riches et beaucoup d’espèces profitent des eaux réchauffées l’été, des fonds riches en sédiments et en débris végétaux ainsi que de la forte végétalisation du cours d’eau et de ses rives. Ailleurs, sur les rivières plus rapides et souvent de dimensions plus modestes, de nombreux ouvrages hydrauliques (barrages, seuils) ont souvent modifié de manière importante le régime naturel du cours d’eau, et des zones calmes, relativement profondes avec des fonds vaseux plantés de nombreux hydrophytes, ont permis aux mêmes cortèges de s’installer. L’espèce caractéristique qui domine ici et qui survole inlassablement ces eaux calmes est l’Anax empereur Anax imperator parfois remplacé par l’Anax napolitain Anax parthenope, qui moins agressif, supporte souvent mal la concurrence avec le premier. Tandis que L. fulva puis O. curtisii, plus tard en saison, patrouillent le long des berges, le Gomphe de Graslin Gomphus graslinii demeure au-dessus des herbiers flottants. B. irene peut profiter des berges les plus boisées. Sur certains secteurs de la Charente et de la Dronne, on peut même rencontrer la très rare Cordulie splendide Macromia splendens. Le cortège caractéristique est aussi composé de nombreuses espèces de zygoptères. Des populations nombreuses de C. splendens sont généralement présentes. Au sud de la région, elles côtoient et parfois se mélangent avec les peuplements du Caloptéryx occitan Calopteryx xanthostoma. Les zones enherbées des rives servent de terrain de chasse aux Platycnémididés, surtout l’Agrion blanchâtre Platycnemis latipes et P. pennipes alors que les nombreux herbiers d’hydrophytes affleurant (Myriophyllum spp. et Ceratophyllum spp. surtout) leur permettent de déposer leur ponte puis d’abriter les larves. Sur l’eau, au-dessus de ces mêmes herbiers, E. lindenii est omniprésent alors que les deux autres espèces d’Erythromma, la Naïade au corps vert E. viridulum et plus rarement la Naïade aux yeux rouges E. najas, marquent une préférence pour les hydrophytes flottants (Nympha lutea ou Lemna spp. notamment). Bien sûr, plusieurs espèces plus ubiquistes ou ayant une écologie plus plastique enrichissent souvent ces cortèges, en particulier C. puella, I. elegans, l’Agrion orangé Platycnemis acutipennis, le Leste vert Chalcolestes viridis, le Gomphe joli Gomphus pulchellus, ou encore O. forcipatus.
Les grandes rivières picto-charentaises sur lesquelles il est possible d’observer ces cortèges sont : le Clain (86), la Vienne (en amont des barrages de l’Isle-Jourdain), la Dronne, le cours aval de la Charente (16 et 17), les cours moyen et aval de la Sèvre niortaise (17 et 79) et la partie aval du Thouet (79). Cependant, de nombreuses autres rivières moyennes offrent, sur certains secteurs, des habitats identiques.





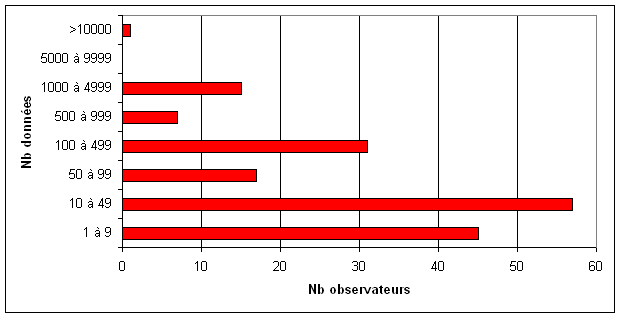
 A l’origine, uniquement prévue pour le groupe odonates de la Vienne, « La Libelluline », créée en 2004, était une publication ouverte à tous les amoureux des Libellules. Il s’agissait d’un outil permettant de communiquer et d’échanger des informations entre naturalistes.
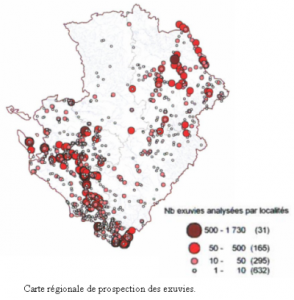
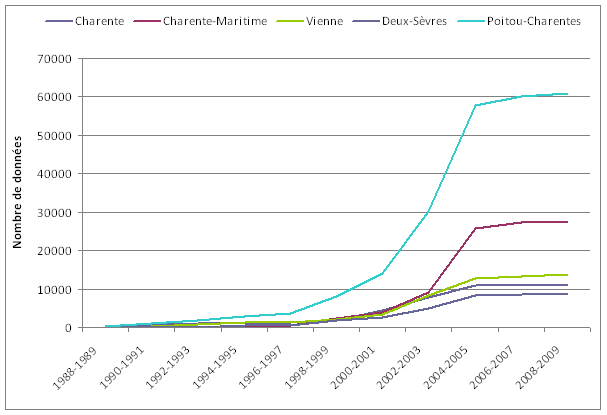
A l’origine, uniquement prévue pour le groupe odonates de la Vienne, « La Libelluline », créée en 2004, était une publication ouverte à tous les amoureux des Libellules. Il s’agissait d’un outil permettant de communiquer et d’échanger des informations entre naturalistes. Réalisé en 2001 dans chaque département de la région par les coordinateurs, cet état des connaissances de l’odonatofaune était basé sur l’analyse des données bibliographiques et des collections privées, ainsi que des données de terrain associatives. Ces analyses croisées utilisant uniquement les informations vérifiables et contrôlées sur un lot de données relativement restreint, ont permis de dresser un premier bilan des espèces présentes en région Poitou-Charentes ainsi que leur répartition. De fortes lacunes sont donc apparues, aussi bien sur le nombre d’espèces recensées dans chaque département, que sur leur répartition spatiale.
Réalisé en 2001 dans chaque département de la région par les coordinateurs, cet état des connaissances de l’odonatofaune était basé sur l’analyse des données bibliographiques et des collections privées, ainsi que des données de terrain associatives. Ces analyses croisées utilisant uniquement les informations vérifiables et contrôlées sur un lot de données relativement restreint, ont permis de dresser un premier bilan des espèces présentes en région Poitou-Charentes ainsi que leur répartition. De fortes lacunes sont donc apparues, aussi bien sur le nombre d’espèces recensées dans chaque département, que sur leur répartition spatiale.